What happens after a living thing eats another living thing? In general, we assume that the one disappears, and the other goes on. In fact, it is seen as an absolutely necessary part of life—the eater breaks down the eaten into its molecular components, derives energy and nutrient building blocks for its own body from the food, and excretes the rest as waste. Such is life in the food chain—some eat others until they themselves succumb to being consumed. Both a scientific and metaphysical model of being, modern notions of nutrition—increasingly described under the term “metabolism” since the mid-nineteenth century—have centered on the principle of inter-conversion, what Thomas Huxley described in 1869 as the “transubstantiation” of one kind of protoplasm to another in the body’s internal chemical laboratories, in which food served to fuel both the engine of the body and the flame of thought; I burn, he lectured, so that others may have light. 1 This version of metabolism has served as one of the defining or delimiting characteristics of life itself, alongside the other universal prerequisite: reproduction. Living things are things that metabolize and reproduce.
Contemporary events in the sciences of metabolism and genetics are beginning to show the historical peculiarity of these longstanding and very stable frameworks for defining metabolism. Destabilization of two central and interlinked features of metabolism is underway. The first is the logic of complete conversion: that food becomes, in the machinery of metabolism, components that are used by the eater to ends irrelevant to the eaten. The dispersion of seeds and the lodging of heavy metals in tissues excepted, the general idea that ingested food is fuel, that it is a substrate fed into a chemical system, that heat and movement and bodily substance are made out of it, and the rest is excreted in the form of waste still seems just like “common sense.” Food is nutrition, it is sustenance, and for humans it might also be allowed to be culture—although that quality is generally seen as fairly irrelevant to its biological function. What else could it be?
A second historical characteristic of metabolism currently coming undone in laboratories and experiments today is the deep trench between metabolism and reproduction or replication. There is slippage between reproduction and replication because the domain of biological reproduction—how cells and multicellular organisms propagate to make more—is in and after twentieth-century life sciences largely subsumed to the genetic, whereby the inheritance of form and function from one living being to its offspring is understood to occur through the mechanisms of DNA replication and cell division. 2 The important functions of gene regulation and replication have been seen as quite distinct from those of metabolism, different topics for different disciplines. 3 This too seems “common sense”: metabolism and reproduction are like two pillars keeping up the same roof, two defining characteristics of life that are separate together. One converts, the other propagates using the energy and materials derived from conversion. They exist in overlapping but essentially separate temporal organizations that map nicely onto germ and soma, itself a distinction arising at the end of the nineteenth century: reproduction concerns the question of how threads of biological substance persist across death and across generations, while metabolism is that which keeps an organism alive day to day, within its lifespan.
Again, we might ask—but what else could it be? Glimpses of quite another mode of incorporation are available today in contemporary biomedical science. Met with manifest surprise by scientists, these emerging narratives show up the historical and cultural particularity of both the logic of conversion and the separation of metabolism from reproduction. This paper begins with an exploration of some very contemporary scientific studies that depict the human body as teeming with genetic components from the plants, microbes, and insects that humans live with and ingest. While commentators have greeted these studies as bearing implications for how we will think about everything from the body to the microbial world to genetically modified food in the future, I am interested in the implications of these studies for the past, for our understanding of how Western modern thought has been organized—or perhaps more accurately, for what has been unthinkable. And subsequently, how this difficulty of thinking, eating, and replicating together is related to structures of comprehension of eating and evolving, eating and thinking, eating and behaving, and eating and species in scientific and cultural theory (including feminist theory) alike. 4 The focus of this paper oscillates—taking interest in theoretical and ontological tumult in present-day life science as an object of inquiry in itself—while asking what the implications of these disturbances are for understanding the history from which they depart.
An Example to Begin With: The Plants in Animals
In September 2011, a group of molecular biologists at Nanjing University, China, published an article in the journal Cell Research claiming to have found plant genetic material in the mouse liver; the plant molecules, they posited, participate in regulating mouse genes that code for proteins involved in blood lipid regulation. This report, “Exogenous plant MIR168a specifically targets mammalian LDLRAP1: evidence of cross-kingdom regulation by microRNA,” found that a plant-derived RNA molecule was entering the blood and tissues of mice and humans after they ate rice. 5 There are many kinds of RNA—the kind in question was a microRNA, a very short sequence of 8-24 nucleotides. Messenger RNA, by contrast, is a much longer string—often hundreds of nucleotides—that is transcribed from the DNA sequence and serves as the template for assembling amino acids into particular proteins.
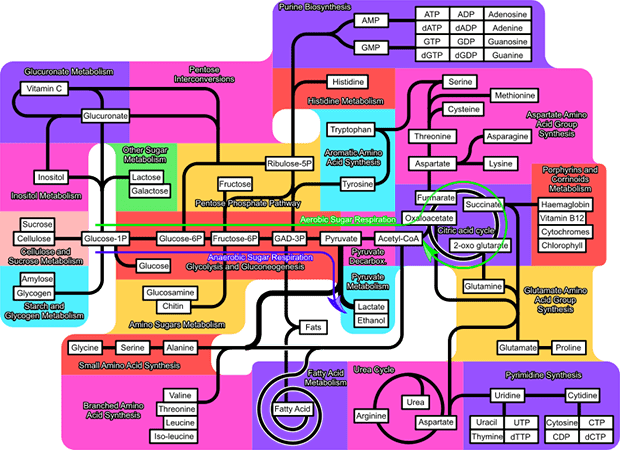
Not only was this small plant microRNA bypassing gastric digestion, its presence and operation in the body was surprising. MicroRNAs “silence” gene expression by sticking to messenger RNA through sequence complementarity. Plant microRNA was found in small lipid bodies (microvesicles) shed by intestinal cells, where it was associated with the protein Argonaute—these complexes were found in the bloodstream of humans and the bloodstream and livers of mice after these experimental subjects had eaten rice. Argonaute is known to be active in the complex of proteins and microRNA that interfere with messenger RNA action—the companionship between the plant microRNA and this particular protein therefore suggested that the plant molecules’ presence was not a random occurrence. Further, the plant microRNA has exact sequence complementarity to a portion of the messenger RNA transcribed by a mammalian gene that codes for a receptor typical to liver cells. The microRNA, because of this sequence complementarity, interrupts the “translation” stage of DNA being transcribed to messenger RNA being translated into protein form. Such a blocking or disintegration of messenger RNA reduces the amount of receptor made by liver cells. In the end, this has the physiological effect of lowering the extraction of low-density lipoprotein (a form of fat) from the bloodstream by the liver.
This report does not make easy reading for the nonscientist. Its gist, however, is clear enough: the research suggests that components of plants enter the bodies that eat them and participate in regulating those bodies’ genes, leading to physiological change (altered blood lipid levels). 6 While it comes as no surprise that the environment of an organism shapes its life in complicated ways, it is actually quite a surprise to find components of plants participating in the general activity of regulation, the determination of whether processes happen or not. This is because food is generally understood to provide the stuff out of which bodies are made, to provide important cofactors and essential nutrients and the energy that bodies need—and is generally not understood to run the operation of being a body.
Responses to this research have ranged from excitement to disbelief. It has been reported that the paper was rejected by other journals for being “too extraordinary” before eventually being published in Cell Research. 7 Other scientists have claimed that the findings are a result of contamination or the mistaking of native animal sequences for exogenous microRNAs—after all, the molecules in question are incredibly tiny. On the other hand, the results begin to look less remarkable as other findings begin to filter out. A group of systems biologists set out to find and sequence all of the microRNAs found in a small number of individuals’ blood plasma, finding to their surprise, “that a significant fraction of the circulating RNA appear to originate from exogenous species.” 8 They found microRNAs in the human bloodstream from bacteria, specifically, from microbes known to live commensally with humans in what has come to be called the gut “microbiome;” they found microRNAs from fungi, including from yeast; they found microRNAs from common food items, with corn, rice, tomato, soybeans, and grapes in the highest numbers; and they found microRNAs from insects, such as honeybees, mosquitoes, and houseflies. It is clear that this is not what they set out to find. The surprised tone of the scientific report is worth quoting here:
It has long been thought that the human body is highly insulated from its environment by two major protective mechanisms, an active and dynamic immune system, and the body’s physical barrier, represented by skin, gut epithelium, mucus membranes, etc. […] While some sequences mapped to known human transcripts and other genomic sequences, we were surprised, even after very stringent screening criteria, to see that a significant proportion of the sequencing reads from human plasma samples in our study clearly originated from various microbes, insects, and food sources. 9
While this study did not elaborate a functional role for these exogenous microRNAs, they did observe that many of these molecules were, as in the other study, associated with lipid vesicles, or the human proteins active in gene regulation. Taken together, this set of findings presents any number of questions, which commentators have generally seen as lying in the future: how many more “foreign” molecules from food will be found to regulate human genes? Will it be the same for other microRNAs, for example those found in the meat humans eat, or is there something biochemically distinct about plant microRNAs that protect them from being digested into their constituent pieces? 10 One commentator has proposed that now “we must also consider nucleic acid ‘information’ as a nutrient,” and therefore we should understand that eating food also means eating information. 11 What are the implications, positive and negative, for genetic modification, food engineering, and edible vaccines? 12 Beneath the carefully remote diction of today’s scientific publication, there is a palpable astonishment at this finding of the activity of plant, microbial, fungal, and insect essences in the human body. Indeed, there is a futuristic cast to these intimations of “inter-kingdom communication,” an idea of plants as deceptively sessile controlling agents, manipulating the animal body at a distance: “Food We Eat Might Control Our Genes” as Scientific American put it. 13 As with the finding that bacteria living commensally in the human gut produce neurotransmitters such as serotonin and gamma-aminobuytric acid (GABA) catecholamines, which immediately makes one rethink the term “neurotransmitter” as a descriptor for this substance; so “information” stutters and fails to capture a phenomenon never imagined by the DNA-as-information model in which the genetic material of a body was in, for, and by that body. 14
It is perhaps less intuitive—seemingly backwards, even—to ask what the implications of these findings are for the past, for the nineteenth and twentieth centuries, rather than for the twenty-first. As much as one can say that it was previously unknown that ingested plant bits and the exudations of diet-derived microbes could regulate animal genes, it was as much unthought. It could be argued, of course, that it is only the technology of RNA identification that has brought us to this point; microRNAs were first characterized in the early 1990s and a widespread regulatory role for them was realized only in the early 2000s. This caused an explosion of interest in the potential pharmaceutical uses of microRNAs, and a hunt for each species’ own microRNAS. It apparently did not occur to anyone to look for plant RNA in animals (or vice versa), since it was assumed that microRNA, like messenger RNA, only arose from the DNA template and circled back to affect the expression of the very DNA from which it came.
The broader point is that one of the controversial surprises of the microRNA findings is the intact functional role of the plant molecule in its passing through the body, a role that is not “cofactor,” “building block” or “energy” provision, nor “drug.” This unnamed functional role—only clumsily captured by the notion of “eating information”—helps makes explicit the unthinkable in previous formulations. Why has it been generally obvious—specifically in Western Euro-American discourse—that plants stop being plants as soon as they are eaten? That the honey does not contain the bee, nor the milk the cow? What is it about the scientific rational model of incorporation that has made it hard to think about the persistence of others in the flesh, the continuing presence of the eaten in the eater? 15
At the same time, this example highlights the profound gap that has existed between metabolism and the DNA sciences. Perhaps in the provision of essential cofactors, food has been seen as a rate-limiting contributor to life processes, but rarely has it been seen to enter into the realm of genetic action, whether regulation or replication. At most, food’s components have been seen as cues from the external environment that can induce gene action after being appropriately “read” by the machinery of gene regulation—they have never been characterized as being themselves directly exerting regulatory power or having genetic action. Such agency for foodstuffs has not been thinkable. The split between the replicative and the metabolic has created the conditions under which scientific findings of their intercalation are experienced as a fundamentally novel discovery: too extraordinary, perhaps, to be true.
With the increasing realization that diet-derived and metabolically-regulated chemical moieties are central to both the regulation and the replication of the genome, it is possible for scientific observers to write that, “the concepts underlying genetics and metabolism are in effect merging […] there is metabolism at all levels of biological organization, from the whole organism, to the cell, and finally down to the genome. 16 There is no genetic action, no manifestation of a genome, without metabolism; nurture metabolizes nature. The instability and rupture in the present moment—the glimpses of an otherwise around the separateness of metabolism and genetics—provides an opportunity to see the same history of biology that we have always had, but to notice in it features that were perhaps always in plain sight and yet not remarkable, exactly because these features were taken to arise from biological matter itself. Thus metabolism and reproduction, and then metabolism and genetics, were taken as given by living matter itself as two essential but essentially separate features of life, organized as a hierarchy in which one function ran the other.
- Thomas Huxley, “The Physical Basis of Life,” The Fortnightly Review 5 (1869): 129-145. Anson Rabinbach, The Human Motor: Energy, Fatigue, and the Origins of Modernity (Berkeley: California UP, 1992). The persistence of this idea that thinking is fueled by nutrition can be seen in the innumerable imaging studies that track glucose consumption in the brain as a proxy for thought.[↑]
- Staffan Müller-Wille and Hans-Jörg Rheinberger, A Cultural History of Heredity (Chicago: Chicago UP, 2012).[↑]
- Scott Gilbert, “Intellectual Traditions in the Life Sciences: Molecular Biology and Biochemistry,” Perspectives in Biology and Medicine 26 (1982): 151-162.[↑]
- Elizabeth Wilson, “Underbelly,” differences 21 (2010): 194-208.[↑]
- Lin Zhang, et al. “Exogenous plant MIR168a specifically targets mammalian LDLRAP1: evidence of cross-kingdom regulation by microRNA,” Cell research 22.1 (2011): 107-126.[↑]
- Only a small percentage of the genome, which is present in every cell in the body, is actually being “expressed” at any given time, going through transcription to messenger RNA and then translation to a protein molecule. The complexity of what is “on” or “off,” or made in high or low quantities in any given cell must be constantly regulated; these processes constitute bodies living in time and in complex changing environments.[↑]
- Emily Willingham, “Plant RNA Paper Questioned,” The Scientist, 16 Apr. 2012.[↑]
- Kai Wang, Hong Li, Yue Yuan, Alton Etheridge, Yong Zhou, David Huang, Paul Wilmes, David Galas. “The Complex RNA Spectra in Human Plasma: An Interface With Human Gut Biota?” PLoS ONE 7.12 (2012): e51009. doi:10.1371/journal.pone.0051009.[↑]
- Wang et al. 2012: 9.[↑]
- Plant microRNAs are methylated, animal microRNAs are not. This appears to protect plant microRNAs from digestion.[↑]
- Hirschi, Kendal D. “New foods for thought,” Trends in plant science 17.3 (2012): 123-125. At the same time, “nutrients” are not generally understood to be direct molecular regulators of genes, so it is a proposal of a new class of substances that has the potential to remake the very idea of a nutrient.[↑]
- Hervé Vaucheret and Yves Chupeau, “Ingested plant miRNAs regulate gene expression in animals,” Cell research 22 (2011): 3-5.[↑]
- Anne-Marie C. Hodge, “Food We Eat Might Control Our Genes,” Scientific American, 5 Nov. 2011. http://www.scientificamerican.com/article.cfm?id=vitamins-minerals-and-microrna[↑]
- David T. Hughes and Vanessa Sperandio, “Inter-kingdom Signalling: Communication Between Bacteria and Their Hosts,” Nature Reviews Microbiology 6 (2008): 111-120.[↑]
- This is not to say that it has been impossible to think outside of this frame; alternative schemas occurred at various points, voicing opposition to elements of the metabolic-historic separation. See, for example, Elizabeth Wilson’s discussion of Sàndor Ferenczi’s theory of phylophagy elaborated in the late 1930s—his idea that the infant suckling is drinking not just from the body of the mother, but of the body of the mother, and her embodiment in turn of her history, and thus the imbibition of the whole history of nutrition, and indeed evolution. Elizabeth Wilson, “Another Neurological Scene.” History of the Present 1.2 (2011): 149-169.[↑]
- Michael L. Power and Jay Schulkin, The Evolution of Obesity (Baltimore: Johns Hopkins UP, 2009): 297-298.[↑]